Cell Fate Protection and Reprogramming


We are applying different genetic screening approaches in order to identify more factors that might play a role in restricting reprogramming of different tissues. For this, we are using a variety of transgenic animals, which allow screening by RNAi and mutagenesis for factors that inhibit direct reprogramming to neuronal or other cell types such as muscle or intestinal cells.
We performed a manual whole-genome RNAi and mutagenesis screen using fluorescence stereo-microscopes to identify mutant phenotypes. This ongoing work yielded a large number of novel factors that appear to antagonize the first step of direct reprogramming, which is the induction of the new fate.


The mutated gene in isolated mutants is identified by using whole genome sequencing (WGS) which is being performed at the BIMSB genomics platform.


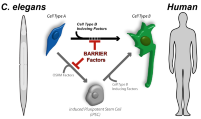
Building our successful validation of a chromatin remodeler as a neuronal reprogramming barrier in both C. elegans and human, we will continue to study whether more of our newly identified factors have conserved functions. Besides chromatin factors also metabolome regulators such as dehydrogenases were identified as reprogramming barriers, which will be further characterized with respect to molecular mechanisms and conservation in mammals. In collaborations with Simone Spuler (Charité) we will continue on elucidating how the human homologs of identified chromatin regulators are implicated in laminopathy using our newly established C. elegans disease models.

Collaborations with Sebastian Diecke (BIH), Magdalena Götz (LMU), and Benedikt Berninger (Mainz University) are ongoing to test reprogramming barriers identified in C. elegans during reprogramming of: 1. human fibroblasts to iPSCs and neurons (Diecke), 2. Astrocytes to neurons (Götz), and 3. Human/Mouse Pericytes to neurons (Berninger).
The detailed characterization of an identified reprogramming factor starts with assessing the temporal and spatial expression pattern in C. elegans. To obtain faithful expression patterns we are using either fosmids (large genomic DNA fragments) to introduce fluorescent proteins by bacterial recombineering into the genomic locus of the gene of interest as described in Tursun et al. 2009 or CRISPR/Cas9-mediated genomic editing. 4D microscopy as well as confocal scanning microscopy can be applied in order to characterize the expression pattern of the gene of interest in living worms.
We can perform tissue-specific chromatin immunoprecipitation (ChIP) in combination with deep-sequencing (ChIP-Seq) to identify genome-wide protein-DNA interactions. Furthermore, purified proteins can be subject to mass spectrometry to identify protein-protein interactions using application such as ‘worm SILAC’.
In the course of characterizing reprogramming factors, we are also interested in assessing changes in chromatin modifications and structure during cell fate specification and conversion. Application of ChIP using in vivo material to elucidate specific histone modifications under certain conditions can be easily applied to C. elegans. Also, our collaboration with the New York University provides us with expertise and support by the Ercan group at NYU. Sevinc Ercan has pioneered applying ChIP to worms and we can benefit from her group’s expertise.



Tursun Beer Hour for BIMSB

Workshop on Metazoan Systems Biology in Bordeaux, October 2018


Congratulations to Ena Kolundzic for defending the PhD Thesis March 2018
Congratulations Stefanie Seelk for defending the PhD Thesis October 2017
The European Worm Meeting (EWM) 2016 took place in Berlin at the MDC!
Congrats to Andreas and Selman for their Prizes at the MDC PhD Retreat 2015!

Congrats to Andreas for winning the Science Slam 2015!

Congrats to Andreas, Ena and Steffi! Prize winners at the MDC PhD Symposium 2014!
